Abstract & Overview
GW-501516 — also designated GW1516, GSK-516, Cardarine, and Endurobol — is a synthetic, orally bioavailable small-molecule agonist of the peroxisome proliferator-activated receptor delta (PPARδ), a ligand-activated nuclear transcription factor that serves as a master regulator of fatty acid catabolism, mitochondrial biogenesis, and skeletal muscle fibre-type programming. Developed through a collaborative research programme between GlaxoSmithKline and Ligand Pharmaceuticals beginning in 1992, GW-501516 was initially investigated as a therapeutic candidate for metabolic syndrome, dyslipidaemia, obesity, and cardiovascular disease [1][2].
The compound displays exceptional receptor selectivity — binding PPARδ with a Ki and EC50 of approximately 1 nM and exhibiting greater than 1,000-fold selectivity over the closely related PPARα and PPARγ subtypes [1]. Preclinical studies demonstrated that GW-501516 shifts skeletal muscle energy substrate utilisation from glucose to fatty acids, increases the proportion of oxidative slow-twitch muscle fibres, enhances running endurance, improves the lipid profile, and protects against diet-induced obesity and type II diabetes in rodent and primate models [3][4][5]. These properties led to its characterisation as an ‘exercise mimetic’ — a compound capable of pharmacologically replicating certain molecular adaptations of endurance training [4].
“PPARβ/δ agonist and exercise training synergistically increase oxidative myofibers and running endurance in adult mice… AMPK-PPARδ pathway can be targeted by orally active drugs to enhance training adaptation or even to increase endurance without exercise.” — Narkar VA et al., Cell (2008) [4].
Despite this compelling preclinical profile, GW-501516 was discontinued by GSK in 2007 following the emergence of carcinogenicity data in animal studies, which demonstrated rapid tumour development across multiple organ systems at research doses. The compound has since been added to the World Anti-Doping Agency (WADA) prohibited list and has never received regulatory approval for human use. It remains an important research tool for elucidating PPARδ biology and the molecular basis of exercise adaptation [6][7].
Molecular Identity and Structural Architecture
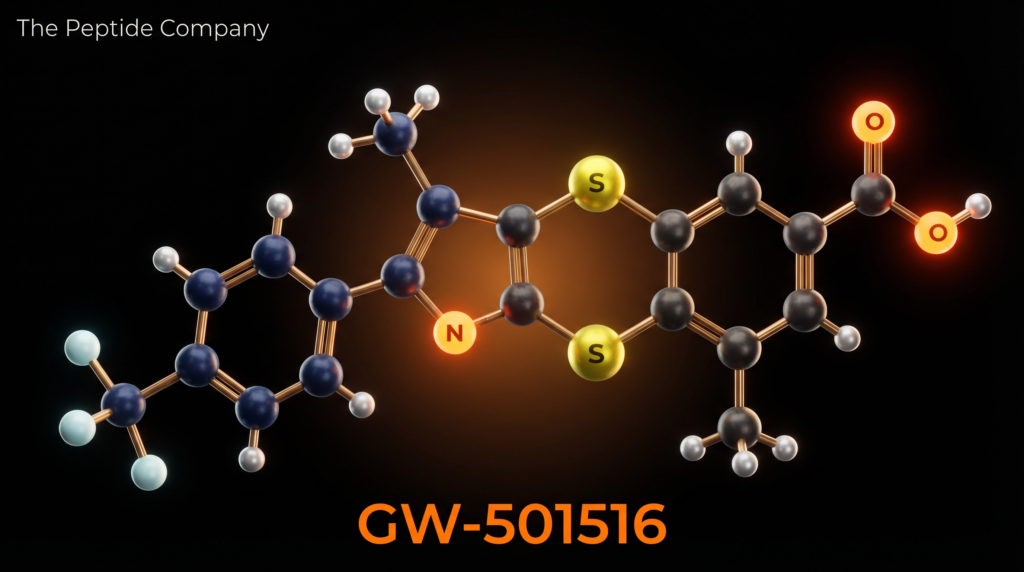
GW-501516 is a small-molecule synthetic compound with the molecular formula C₂₁H₁₈F₃NO₃S₂ and a molar mass of 453.49 g/mol (CAS: 317318-70-0; PubChem CID: 9803963; DrugBank: DB05416). Its IUPAC name is {4-[({4-methyl-2-[4-(trifluoromethyl)phenyl]-1,3-thiazol-5-yl}methyl)sulfanyl]-2-methylphenoxy}acetic acid. The molecule is administered orally and was developed using combinatorial chemistry and structure-based drug design approaches, as reported by Oliver et al. in PNAS (2001) [1].
The structural architecture of GW-501516 comprises four key pharmacophoric elements: a 4-(trifluoromethyl)phenyl group that anchors the molecule within the PPARδ ligand-binding domain; a 4-methyl-1,3-thiazol-5-yl heterocyclic core that provides the scaffold for receptor engagement; a thioether (methylsulfanyl) linker connecting the thiazole to the phenoxy moiety; and a 2-methylphenoxyacetic acid terminus that contributes to binding affinity and metabolic stability. The trifluoromethyl group is particularly important for selectivity, as it creates hydrophobic contacts within the PPARδ binding pocket that are not accommodated by PPARα or PPARγ [1][2].
The compound’s selectivity profile is exceptional: Ki = 1 nM and EC50 = 1 nM for PPARδ, with greater than 1,000-fold selectivity over PPARα and PPARγ. At higher concentrations, some PPARα agonism has been reported, which may contribute to additional lipid-lowering effects via hepatic fatty acid oxidation pathways. GW-501516 is orally bioavailable and has been used extensively as a research tool to dissect the physiological and pathophysiological functions of PPARδ in metabolic disease models [1][2].
Mechanistic Rationale: PPARδ Activation and Downstream Signalling
PPARδ Ligand Binding and PGC-1α Coactivator Recruitment
Upon oral administration, GW-501516 enters the systemic circulation and gains access to target tissues — principally skeletal muscle, adipose tissue, liver, and the cardiovascular system. Within these tissues, GW-501516 binds to the ligand-binding domain of PPARδ with nanomolar affinity, inducing a conformational change in the receptor that facilitates recruitment of the transcriptional coactivator peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α). The resulting PPARδ/PGC-1α complex translocates to the nucleus and binds to peroxisome proliferator response elements (PPREs) in the promoter regions of target genes, initiating a coordinated transcriptional programme centred on fatty acid catabolism and mitochondrial biogenesis [3][4].
Key transcriptional targets of the GW-501516-activated PPARδ/PGC-1α complex include: carnitine palmitoyltransferase 1 (CPT1), the rate-limiting enzyme for mitochondrial fatty acid import; fatty acid binding protein 3 (FABP3), which facilitates intracellular fatty acid transport; pyruvate dehydrogenase kinase 4 (PDK4), which phosphorylates and inactivates the pyruvate dehydrogenase complex, thereby reducing glucose oxidation and sparing glucose for other tissues; uncoupling protein 3 (UCP3), which promotes mitochondrial uncoupling and thermogenesis; and a suite of slow-twitch contractile protein genes including myosin heavy chain I (MHC-I) and slow-isoform troponin I [3][5].
Skeletal Muscle Fibre-Type Switching and Endurance Enhancement
One of the most striking effects of GW-501516 in preclinical models is its capacity to reprogram skeletal muscle fibre composition toward a more oxidative phenotype. Skeletal muscle fibres are broadly classified as type I (slow-twitch, oxidative, fatigue-resistant), type IIa (fast-twitch, mixed oxidative-glycolytic), and type IIb (fast-twitch, glycolytic, fatigue-prone). Endurance exercise training progressively shifts the fibre-type distribution toward type I and IIa, enhancing oxidative capacity and fatigue resistance. GW-501516 pharmacologically mimics this adaptation: Chen et al. (2015) demonstrated that three weeks of GW-501516 treatment increased the proportion of succinate dehydrogenase (SDH)-positive oxidative fibres by 72% in sedentary mice and by 113% in trained mice compared to untreated sedentary controls [3].
The landmark study by Narkar et al. (Cell, 2008) established that GW-501516 and the AMP-mimetic AICAR (an AMPK activator) act synergistically to enhance running endurance. When combined with exercise training, GW-501516 produced near-doubling of running endurance in adult mice. Critically, the study demonstrated that the AMPK-PPARδ axis could be pharmacologically activated to replicate exercise adaptations even in sedentary animals — a finding that generated significant scientific and public interest in the compound as an ‘exercise pill’ [4].
Fatty Acid Oxidation and Glucose Sparing
The metabolomic study by Chen et al. (2015) provided detailed insight into the substrate utilisation shifts induced by GW-501516. Using two-dimensional gas chromatography time-of-flight mass spectrometry (GC×GC-TOFMS), the investigators demonstrated that GW-501516-treated mice preferentially metabolised fatty acids as their primary energy source during exhaustive running, resulting in significantly reduced glucose consumption and lactate formation compared to untreated controls. This glucose-sparing effect — mediated principally through PDK4 upregulation and consequent inhibition of pyruvate dehydrogenase — preserves glycogen stores and delays the onset of fatigue, providing a mechanistic basis for the observed endurance enhancement [3][5].
Fan et al. (Cell Metabolism, 2017) further refined this model, demonstrating that PPARδ activation promotes running endurance specifically by preserving glucose availability in working muscle, rather than solely by increasing fat oxidation capacity. This glucose-sparing mechanism is distinct from the primary fuel-switching effect and suggests that GW-501516 acts through multiple complementary mechanisms to enhance endurance performance [5].
Lipid Metabolism and Cardiovascular Effects
Beyond skeletal muscle, GW-501516 exerts significant effects on systemic lipid metabolism. Oliver et al. (PNAS, 2001) demonstrated that GW-501516 promotes reverse cholesterol transport — the process by which peripheral cholesterol is transported back to the liver for excretion. In obese rhesus monkeys, GW-501516 treatment significantly increased high-density lipoprotein (HDL) cholesterol while reducing very-low-density lipoprotein (VLDL) and triglyceride levels, a lipid profile modification considered cardioprotective [1]. Barroso et al. (Endocrinology, 2011) demonstrated that GW-501516 prevents the high-fat diet-induced downregulation of hepatic AMPK and amplifies the PGC-1α-Lipin 1-PPARα pathway, reducing hepatic lipid accumulation and markers of non-alcoholic fatty liver disease [6].
Research Applications and Experimental Evidence
Metabolic Syndrome and Obesity Models
GW-501516 has been extensively studied in rodent and primate models of metabolic syndrome. In diet-induced obesity models, GW-501516 treatment prevented excessive fat accumulation in both brown adipose tissue (BAT) and white adipose tissue (WAT), improved insulin sensitivity, and reduced fasting glucose levels. The compound’s ability to simultaneously address dyslipidaemia, insulin resistance, and adiposity — the cardinal features of metabolic syndrome — made it an attractive therapeutic candidate prior to the emergence of carcinogenicity concerns [2][8].
Exercise Mimicry and Muscle Adaptation Research
GW-501516 has become an indispensable research tool for studying the molecular mechanisms of exercise adaptation. Its ability to pharmacologically activate the PPARδ transcriptional programme has allowed researchers to dissect the contributions of specific metabolic pathways to endurance performance, independent of the confounding variables associated with exercise training protocols. Studies using GW-501516 have elucidated the roles of CPT1-mediated fatty acid import, PDK4-mediated glucose sparing, and PGC-1α-driven mitochondrial biogenesis in exercise-induced muscle remodelling [3][4].
Anti-Inflammatory and Hepatoprotective Research
PPARδ activation by GW-501516 has demonstrated anti-inflammatory properties in several preclinical models, including suppression of NF-κB-mediated inflammatory signalling and reduction of macrophage-derived pro-inflammatory cytokines. In hepatic models, GW-501516 reduced markers of non-alcoholic steatohepatitis (NASH) and improved hepatic lipid metabolism. These findings have positioned PPARδ agonism as a potential research avenue for inflammatory liver disease, though the carcinogenicity profile of GW-501516 itself precludes its clinical development [6][8].
GW-501516 vs. Other PPARδ/Metabolic Modulators: Comparative Profile
| Parameter | GW-501516 (Cardarine) | GW0742 (PPARδ) | SR9009 (Rev-erb) |
| Primary Target | PPARδ (Ki = 1 nM) | PPARδ (EC50 ~1 nM) | Rev-erbα/β |
| Selectivity | >1000-fold over PPARα/γ | High PPARδ selectivity | Rev-erb selective |
| Endurance Effect | Yes (strong, preclinical) | Yes (moderate) | Yes (circadian) |
| Lipid Profile | ↑ HDL, ↓ VLDL/TG | ↑ HDL, ↓ TG | ↓ TG, ↓ cholesterol |
| Carcinogenicity | Yes (multi-organ, rodents) | Limited data | Not established |
| Clinical Status | Discontinued (2007) | Research tool only | Research tool only |
Safety Profile and Regulatory Status
The safety profile of GW-501516 is defined primarily by its carcinogenicity findings in preclinical animal studies. GSK’s internal carcinogenicity studies, presented at the 2009 Society of Toxicology Annual Meeting by Geiger et al. and Newsholme et al., demonstrated that GW-501516 caused rapid tumour development in multiple organ systems — including liver, stomach, tongue, skin, bladder, ovaries, uterus, and testes — in both rats and mice at doses of 3 mg/kg/day [7]. The speed and multi-organ nature of tumour induction was described as unprecedented and led to the immediate discontinuation of the development programme in 2007 [8].
The proposed mechanism of carcinogenicity is paradoxically the same as the mechanism of metabolic benefit: PPARδ activation promotes cell proliferation and reduces apoptosis as part of its pro-survival transcriptional programme. In normal metabolically active tissues such as skeletal muscle, this anti-apoptotic, pro-proliferative effect is beneficial. However, in pre-neoplastic or initiated cells, the same signalling programme accelerates tumour progression. A 2018 study specifically demonstrated that GW-501516 enhanced the growth of colitis-associated colorectal cancer in mice by increasing inflammation and upregulating the glucose transporters GLUT1 and SLC1A5, providing a molecular mechanism for the pro-tumorigenic effect [9].
In 2013, WADA took the unusual step of issuing a public warning to athletes, stating that ‘clinical approval has not, and will not be given for this substance.’ GW-501516 was added to the WADA prohibited list in 2009 under the category of ‘hormone and metabolic modulators.’ Australia classified it as a Schedule 9 prohibited substance in June 2018. Multiple professional athletes across cycling, athletics, and boxing have received suspensions following positive tests for GW-501516 [7][10].
Conclusion
GW-501516 (Cardarine) represents one of the most pharmacologically potent and mechanistically well-characterised PPARδ agonists ever developed. Its capacity to reprogram skeletal muscle metabolism toward fatty acid oxidation, drive fibre-type switching from glycolytic to oxidative phenotypes, enhance endurance performance, and improve the systemic lipid profile established it as a compelling research compound and potential therapeutic candidate for metabolic syndrome. The landmark studies by Oliver et al. (2001), Narkar et al. (2008), and Chen et al. (2015) collectively demonstrated that the AMPK-PPARδ signalling axis is a tractable pharmacological target for mimicking or potentiating the molecular adaptations of endurance exercise.
However, the rapid multi-organ carcinogenicity observed in preclinical studies at research doses represents a fundamental obstacle to clinical translation. The same PPARδ-driven pro-proliferative and anti-apoptotic programme that confers metabolic benefits in healthy muscle tissue appears to accelerate tumour progression in pre-neoplastic cells, creating an inherent therapeutic liability. GW-501516 therefore occupies a unique position in pharmacological research: a compound of extraordinary scientific value for understanding exercise biology and metabolic regulation, whose clinical development was foreclosed by an unacceptable safety profile. Future research in this space will likely focus on identifying tissue-selective PPARδ modulators that retain metabolic benefits while dissociating the pro-tumorigenic transcriptional programme.
References
[1] Oliver WR, Shenk JL, Snaith MR, et al. A selective peroxisome proliferator-activated receptor delta agonist promotes reverse cholesterol transport. Proc Natl Acad Sci USA. 2001;98(9):5306–11. doi:10.1073/pnas.091021198. PMID: 11309497.
[2] Dressel U, Allen TL, Pippal JB, et al. The peroxisome proliferator-activated receptor beta/delta agonist, GW501516, regulates the expression of genes involved in lipid catabolism and energy uncoupling in skeletal muscle cells. Mol Endocrinol. 2003;17(12):2477–93. doi:10.1210/me.2003-0151. PMID: 14525954.
[3] Chen W, Gao R, Xie X, et al. A metabolomic study of the PPARδ agonist GW501516 for enhancing running endurance in Kunming mice. Sci Rep. 2015;5:9884. doi:10.1038/srep09884. PMID: 25943561.
[4] Narkar VA, Downes M, Yu RT, et al. AMPK and PPARδ agonists are exercise mimetics. Cell. 2008;134(3):405–15. doi:10.1016/j.cell.2008.06.051. PMID: 18674809.
[5] Fan W, Waizenegger W, Lin CS, et al. PPARδ promotes running endurance by preserving glucose. Cell Metab. 2017;25(5):1186–1193.e4. doi:10.1016/j.cmet.2017.04.006. PMID: 28467934.
[6] Barroso E, Rodríguez-Calvo R, Serrano-Marco L, et al. The PPARβ/δ activator GW501516 prevents the down-regulation of AMPK caused by a high-fat diet in liver and amplifies the PGC-1α-Lipin 1-PPARα pathway. Endocrinology. 2011;152(5):1848–59. doi:10.1210/en.2010-1468. PMID: 21363937.
[7] Geiger LE, Dunsford WS, Lewis DJ, et al. Rat carcinogenicity study with GW501516, a PPAR delta agonist. 48th Annual Meeting of the Society of Toxicology. Baltimore. 2009.
[8] Sahebkar A, Chew GT, Watts GF. New peroxisome proliferator-activated receptor agonists: potential treatments for atherogenic dyslipidemia and non-alcoholic fatty liver disease. Expert Opin Pharmacother. 2014;15(4):493–503. doi:10.1517/14656566.2014.876992. PMID: 24428677.
[9] Luo Y, Yang Z, Su L, et al. Non-cancerous PTPRO expression is associated with colitis-associated colorectal cancer. Cell Physiol Biochem. 2018;47(6):2472–2484. doi:10.1159/000491627.
[10] Park J, Kim JY. Cardarine (GW501516) effects on improving metabolic syndrome. J Health Sports Kinesiol. 2021;2(2):22–27. doi:10.47544/johsk.2021.2.2.22.
[11] Barish GD, Narkar VA, Evans RM. PPAR delta: a dagger in the heart of the metabolic syndrome. J Clin Invest. 2006;116(3):590–7. doi:10.1172/JCI27955. PMID: 16511591.
[12] Weihrauch M, Handschin C. Pharmacological targeting of exercise adaptations in skeletal muscle: benefits and pitfalls. Biochem Pharmacol. 2018;147:211–220. doi:10.1016/j.bcp.2017.10.006. PMID: 29061342.
Disclaimer: This article is intended strictly for research and educational review purposes. GW-501516 (Cardarine) is a discontinued drug candidate that has never received regulatory approval for human use. It has been shown to cause rapid multi-organ carcinogenicity in animal studies and is classified as a prohibited substance by WADA. This document does not constitute medical advice, endorsement of any substance, or guidance for personal use. All referenced studies were conducted in preclinical (in vitro or animal) models unless otherwise stated.
thepeptidecompany.xyz | Research Division
GW-501516 is studied as a PPAR-δ (PPAR-delta) agonist and its role in regulating lipid metabolism, fatty acid oxidation, and energy utilization in experimental models.
It activates PPAR-δ receptors, which are involved in gene expression related to fatty acid transport, mitochondrial activity, and energy expenditure.
Research focuses on pathways including lipid oxidation, glucose metabolism, skeletal muscle energy utilization, and mitochondrial biogenesis.
PPAR-δ activation is associated with enhanced oxidative metabolism in muscle tissue, making it a target of interest in studies on energy efficiency and endurance pathways.
No, it is a synthetic small-molecule compound, not a peptide, but is often grouped with metabolic research compounds due to its pathway effects.
Some long-term animal studies have reported adverse findings, which is why it is strictly limited to controlled research settings and not approved for human use.
PMID:
12471285 — PPAR-δ activation and lipid metabolism regulation
15102836 — GW-501516 and fatty acid oxidation pathways
16407105 — PPAR-δ agonists and skeletal muscle metabolism
16849545 — Mitochondrial function and oxidative metabolism via PPAR-δ
18316364 — GW-501516 and endurance-related metabolic pathways
19752120 — PPAR-δ role in glucose and lipid homeostasis
20631354 — Gene expression changes induced by PPAR-δ activation
22955512 — Long-term effects and safety findings of GW-501516
AICAR : AMPK Activation, Cellular Energy Sensing, and Exercise‑Mimetic Signaling in Research Models
BAM15 — Mitochondrial Uncoupler Research Article (Educational • Research Use Only)
MOTS-c: The Mitochondrial-Encoded Peptide for Metabolic Regulation and Cellular Resilience
Semaglutide : GLP-1 Receptor Agonism, Incretin Signaling, and Metabolic Regulation
Orforglipron : Oral Small-Molecule GLP-1 Receptor Agonist and Incretin Pathway Modulation
